
https://chenglab.org/about
Contact
- hjcheng@gate.sinica.edu.tw
- (L) 886-2-2789-9971
- 223, Institute of Molecular Biology, Academia Sinica
Research
Axonal Connection, Hippocampus, and Aging
(A) Current Research:
Our current research focuses on how aging processes change the new axonal connections in adult hippocampus. In adult mammalian brain, the subgranular zone of the hippocampal dentate gyrus (DG) retains the capacity for neurogenesis throughout life. These adult-born neurons are functionally integrated into existing circuitry, where they are involved in learning and memory and disruptions to such adult hippocampal neurogenesis (AHN) is implicated in human neurological disorders, including Alzheimer’s disease, depression, epilepsy, and age-related cognitive decline.
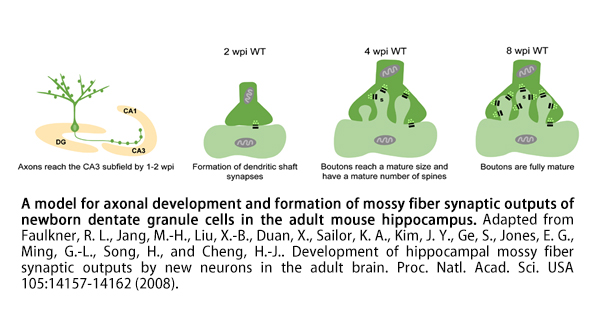
Our objective is to understand why AHN becomes less efficient in the aged brain. Like their mature counterparts, newborn dentate granule cells (GCs) in the adult brain receive synaptic inputs from the perforant pathway and send axonal projections along the mossy fiber pathway into stratum lucidum of the CA3 subfield. In CA3, the newborn mossy fiber boutons (MFBs) establish synaptic contacts with large spiny complexes termed thorny excrescences at the proximal dendrites of mature CA3 pyramidal neurons. We and others have shown that it takes about eight weeks for newborn GCs in young adult mouse hippocampus to form mature MFB contacts with CA3 pyramidal neurons. Adult-born MFBs establish novel synaptic contacts in two main patterns: 1) de novo synaptogenesis - nascent synaptic contacts are formed and mature over time and 2) replacement of preexisting contacts through a progressive encroachment on postsynaptic territory. The mechanisms underlying such development are unknown.
We combine mouse genetics, confocal and electron microscopic imaging, and array tomography-based molecular phenotyping to delineate how newborn GCs change with age and elucidate the underlying mechanisms that control the generation and synaptic integration of newborn GCs in aged mice. We have been using an inducible genetic labeling technique that allows us to control the number of labeled newborn neurons at any age throughout adult life. This mouse line (GliCreER) carries the tamoxifen-inducible CreER protein under the control of a Sonic hedgehog-responsive Gli1 promoter element, which is specifically activated in neural precursor cells. When crossed to reporter mouse lines, GliCreER mice express fluorescent markers only in progenitors and newborn neurons in the DG. The number of labeled neurons can be controlled by the amount of tamoxifen that is injected.
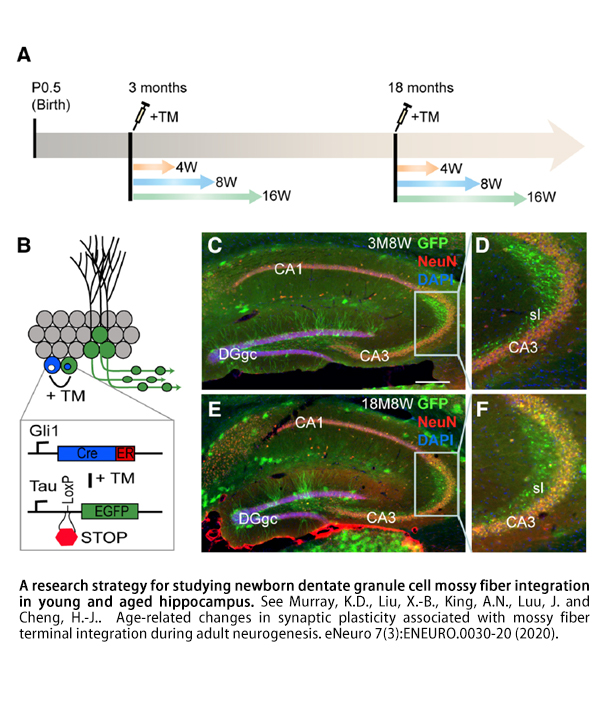
Our current working hypothesis is that the developmental origin, molecular identity, cellular differentiation, and synaptic integration of neuronal progenitors are significantly changed or compromised during the aging process. We plan to systematically determine the molecular and cellular changes criticial for establishing synaptic integration during AHN in young and aged brain. Despite a rich knowledge of the roles of molecular, cellular, and neural activities in regulating AHN, the reasons why neurogenesis is less efficient in the aged hippocampus are still poorly defined. We hope to identify the mechanisms that regulate the formation of newborn GCs in the aged hippocampus, thereby filling a major gap in our current understanding of neurogenesis in the aged brain.
(B) Previous Research Contributions:
(I) Our early research work laid the foundation for the use of AP (alkaline phosphatase) fusion protein techniques and the molecular mechanisms of topographic map in the visual system. Prior to our early research, Ephs were orphan receptors without known functions. We developed the AP fusion expression cloning techniques and identified ephrins. We then developed the RAP (receptor AP) and LAP (Ligand AP) to demonstrate Ephs and ephrins are the “Sperry” molecules for establishing the visual topographic map. Both the AP fusion technique and the biology of Ephs and ephrins in the formation of topographic neural circuits have contributed significantly to the progress of the field.
(II) Using mouse genetics, we generated plexin mutant mice to address the in vivo functions of semaphorin-plexin signaling in axon guidance. These studies demonstrated the differential roles of plexin family members in guiding axons in vivo, and unexpectedly revealed the role of semaphorin-plexin signaling in regulating stereotyped axon pruning in the central nervous system.
(III) To understand the signaling pathways that regulate axon repulsion, we turned to C. elegans genetics and isolated max (motor neuron axon guidance) mutants. We identified and cloned max-1 and max-2. We demonstrated that MAX-1 is involved in AP3-mediated trafficking and degradation of UNC-5 receptor through SUMOylation regulation. We also showed that MAX-2 is a major downstream effector of RAC GTPases that are required for the repulsion of motor axons. These studies provided a foundation for investigating the role of the schizophrenia-related gene disc1 in neural development. By combining C. elegans and mouse genetics, we revealed the previously unknown function of disc1 in regulation of adult-generated neurons in the hippocampus and established a heterologous transgenic disc1 line in C. elegans.
(IV) Axon pruning in the brain had been recognized as a fundamental process for developmental plasticity, but the understanding of the molecular and cellular regulations for this process had been challenging. Focusing on the pruning events in hippocampal and visual circuits, we developed immuno-electron microscopic and imaging analysis to investigate the cellular mechanisms. These studies demonstrated that the axon branches to be pruned often form synapses, suggesting the importance of neural activity for the process.
(V) Using ferret as a model, we studied the role of neural activity in refining the anatomical and functional connections of the visual system. We had identified activity-dependent axon-axon competition as an important mechanism for axon terminal targeting during development and revealed the role of stage III retinal waves in promoting circuit refinement.